Please note that this material is copyright to the publishers of Coral Reefs and to the authors
Not to be reproduced in any way without permission except for personal study

Coral Reefs (1988) 6: 195-205
Studies on Halimeda I.
The distribution and species composition of Halimeda meadows throughout the Great Barrier Reef Province *
Edward A. Drew and Kay M. Abel
Australian Institute of Marine Science, P. M. B. No. 3, Townsville M.S.O., Oueensland 4810, Australia
* Contribution No. 367 from the Australian Institute of Marine Science
Accepted 3 September 1987
GO TO Title-
Abstract-
Introduction-
Methods-
Results-
Discussion-
References
Abstract
Large areas of the inter-reefal seabed in the Great Barrier Reef are carpeted with vegetation composed almost entirely of the green calcareous alga Halimeda. These meadows occur principally in the northern sections between 11o 30' and 15o 15'S at depths of 20 to 40 m, but there are also some in the central and southern sections where they have been found at depths down to 96 m. The vegetation is dominated by the same sprawling Halimeda species that are common on coral reefs in this region. However, on reefs these species grow on solid substrata, not soft sediments like the Halimeda-rich gravels that underlie the meadows. A total of 12 Halimeda species, together with two Udotea and one Penicillus species, are characteristic components of the shallow meadows. Below 50 m depth, species composition is restricted to only two major components. One, H. copiosa, is also important shallower, but the other is an unusually large and heavily calcified form of H. fragilis, a species that is normally a minor, fragile component of the shallow meadows. The maximum biomass found in these meadows was 4637 gm2 of calcareous algae, although the mean for vegetated areas was 525 gm2 . These meadows are confined to the nutrient-depleted waters of the outer continental shelf just inside the outer barrier reefs, and are usually associated with distinct shoaling of the seabed caused by accumulation of thick deposits of calcareous Halimeda segments. The meadows are probably supported by very localized upwelling of nutrients from the adjacent Coral Sea onto the shelf, where they enrich the otherwise nutrient-depleted waters.
GO TO Title-
Abstract-
Introduction-
Methods-
Results-
Discussion-
References
Introduction
Halimeda, a calcified green alga of warm seas, often grows well on corals reefs either in sand, anchored by rhizoidal holdfasts, or on solid substrata. In addition to considerable primary productivity by living Halimeda (Hillis-Colinvaux 1980), the aragonitic segments of dead plants contribute substantially to reefal sediments (Merten 1971; Wiman and McKendree 1975; Neuman and Land 1975; Drew 1983).
In a detailed study of the distribution and growth rates of Halimeda growing on reefs in the central region of the Great Barrier Reef (GBR), Drew (1983) mentioned a total of 16 species and provided quantitative data on the distribution of the 13 lithophytic representatives. That study showed Halimeda opuntia and a taxon then considered to be H. copiosa were co-dominant at all sites studied on the continental shelf, but on a nearby oceanic plateau reef in the Coral Sea H. minima and H. distorta dominated the Halimeda biomass, which was considerably greater than found on the shelf reefs.
Maxwell (1968, 1973) first reported extensive deposits of Halimeda-rich gravels away from reefs, on the inter-reefal seabed in the GBR, particularly in the very south and the north of the Province. Similar deposits have since been reported around Lizard Island (Orme et al. 1978; Orme 1985), Great Detached Reef (Hopley 1978), and the Ribbon Reefs off Cooktown (Davies and Marshall 1985; Phipps et al. 1985). All these geological studies have mentioned luxuriant algal communities, usually dominated by Halimeda, growing on the inter-reefal gravels.
Such accumulations of porous carbonate sediments are very reminiscent of the phylloid algal bioherms that occur at shelf breaks in late Paleozoic reef systems and that may now be acting as hydrocarbon reservoirs (Ball et al. 1977; Wray 1977). Further information on the factors controlling their rates of formation and geographical distribution is therefore of considerable importance.
We have extended our reefal Halimeda studies to these inter-reefal areas of the outer continental shelf of the GBR. They are indeed clothed by luxuriant meadows of algal vegetation dominated by Halimeda (Drew and Abel 1985), and this paper describes that inter-reefal community in detail and also its spatial distribution in relation to the associated thick deposits of Halimeda-rich gravels.
GO TO Title-
Abstract-
Introduction-
Methods-
Results-
Discussion-
References
Methods
The entire 1850 km length of the GBR Province is a Marine National Park and we have used its major sections, set out in Fig. 1, as a primary geographical basis in this paper. However, these sections, particularly in the Far North, contain vast areas of inter-reefal Halimeda meadows; so we have further subdivided them into geographically discrete localities, each spanning about half a degree of latitude and named after a significant feature therein. These are also shown in Fig. 1. We use the term Halimeda MEADOW to describe inter-reefal areas of luxuriant Halimeda vegetation, Halimeda MOUND to describe the discrete circular mounds of Halimeda debris that accumulate under these meadows and are of the order of 250 m across and 10 m high, and Halimeda BANK to describe shoaling areas of seabed that occupy several square kilometres and are comprised of numerous Halimeda mounds that sometimes coalesce into ridges.
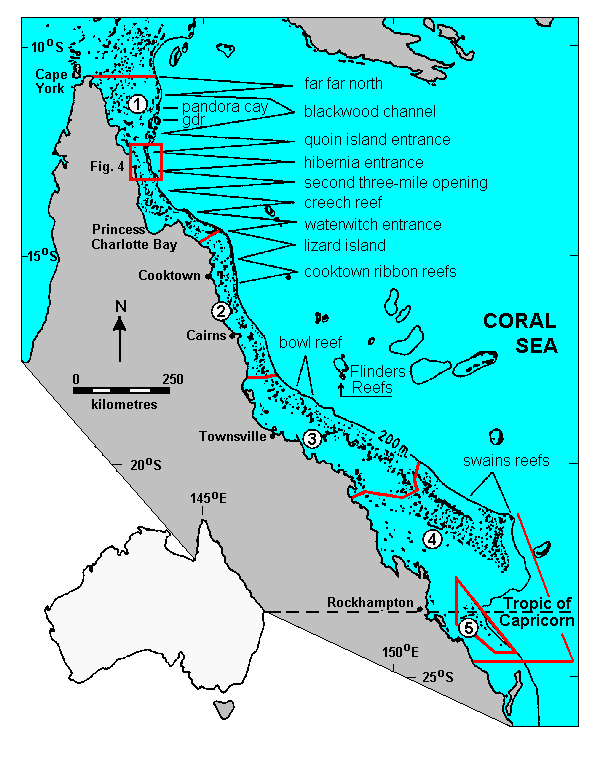
|
|
Fig. 1. The study area, showing the major sections recognized within the GBR, the localities studied, and the position of the map in Fig. 4. Sections (local names in parentheses) 1 Far North; 2 Northern (Cairns and Cormorant Pass); 3 Central; 4 Southern (Capricorn); 5 Far South (Capricornia). Locality names as indicated; gdr = Great Detached Reef
|
Algal communities were sampled quantitatively using grab sampling and Scuba diving techniques. A total of 1013 Van Veen grab samples, each comprising all the benthos from 0.1 m2 of seabed and the underlying sediment to a depth of 0.1 m, were obtained at 281 sampling stations. Up to five samples were taken at most stations, but only one or two were taken when the sediment was pure mud and the station obviously not on a Halimeda meadow. Accurate sampling depths, free from ambiguities arising from steep wire angles and echogram interpretation, were obtained by attaching to grabs a Scuba diver's depth gauge equipped with a maximum depth indicator. At six of the 13 localities, ( lantitative biomass samples were also collected from at least one station by Scuba divers using the methods described by Drew (1983) to sample belt transects 0.25 m wide and several metres long. A total of 76 diver-collected biomass samples were obtained thus from 12 stations.
Living plants of Halimeda, Udotea and Penicillus from both grab and diver samples were air-dried on board ship and then sorted in the laboratory into species, oven-dried at 80 oC and weighed. The organic matter content of Halimeda material was determined from weight loss after combustion overnight at 450 oC in a muffle furnace. Dried calcareous algae, samples of the accompanying sediment and formaldehyde-preserved material of other plants from the grab samples have been retained. These, together with larger quantities of plants obtained from 10-min benthic dredge hauls taken at a number of stations, are available for further study.
GO TO Title-
Abstract-
Introduction-
Methods-
Results-
Discussion-
References
RESULTS
Halimeda species--
Species composition--
Species diversity vs depth--
Biomass--
Other algae/seagrasses--
Spatial distribution
Results
The Halimeda species involved
The taxonomy of the genus Halimeda, described in detail by Hillis-Colinvaux (1980), is based to a large extent on microscopic features. In view of the large amount of Halimeda biomass to be sorted to the species level in this study, we have used macroscopic characters supplemented only by a x 30 hand magnifier and requiring no manipulation beyond snapping segments in half to view cross-sections of skeletal structure. Using these methods we can recognize at least 19 taxa of Halimeda within the GBR Province and have diverged from Hillis Colinvaux's 1980 classification scheme in only one respect, the resurrection of her species H. hederacea (Barton) n. comb. (Hillis-Colinvaux 1968), which she subsequently merged (Hillis-Colinvaux 1969) with H. copiosa, then newly described from the Caribbean by Goreau and Graham (1967). Drew and Abel (1985) used the valid, but now clearly inappropriate, name H. opuntia var hederacea to distinguish such material from H. copiosa, which it was initially called by Drew (1983) at Professor Hillis-Colinvaux's suggestion. However, material much more typical of the Caribbean H. copiosa, albeit with somewhat smaller segments, grows interspersed with the former taxon throughout the GBR. The consistently greater segment size, distinctly angular and often auriculate segment shape, and much greater size of surface utricles of H. hederacea seem sufficient to distinguish it from both typical H. copiosa and H. opuntia at the species level, while differences in gametangial structure suggest closer affinity with H. copiosa/ than H. opuntia. All three taxa maintained their integrity when plants grew vegetatively for many months under identical conditions in the open-circuit aquarium system described by Drew and Abel (1988).
RESULTS
Halimeda species--
Species composition--
Species diversity vs depth--
Biomass--
Other algae/seagrasses--
Spatial distribution
Species composition - calcareous green algae
The species composition of calcareous green algae, mainly Halimeda species, from the inter-reefal Halimeda meadows at all the localities studied is set out in Tables 1 a and b, and also included is a comparison with that of reefal Halimeda vegetation. Little is known of Halimeda on reefs in the northern GBR, so species are arranged in the order of their quantitative importance on reefs in the central GBR.
Table la. Species composition of Halimeda meadows compared with Halimeda vegetation on reefs in the GBR.
Values expressed as % total Halimeda, Udotea and Penicillus biomass in those grab samples which containing at least some Halimeda.
Reefal data from Drew (1983)
|
| Species |
Meadows |
Central GBR
reefs |
Flinders
(oceanic) a
|
|
| H. opuntia (L.) Lamouroux
| 0.6 | 40.2 | 11.6 |
| H. hederacea (Barton) Colinvaux
| 48.4 | 38.3 b | - |
| H. copiosa Goreau and Graham
| 26.0 | 2.5 b | 0.5 |
| H. minima (Taylor) Colinvaux
| - | 0.2 | 55.3 |
| H. micronesica Yamada
| 0.6 | 10.2 | 2.8 |
| H. distorta (Yamada) Colinvaux
| 4.4 | - | 20.8 |
| H. melanesica Valet
| - | 3.8 | - |
| H. lacunalis Taylor
| 0.5 | 1.0 | 4.1 |
| H. macrophysa Askenasy
| - | 2.1 | - |
| H. fragilis Taylor
| 5.6 | t | 4.3 |
| H. tuna (Ellis and Solander) Lamouroux
| 2.0 | 1.0 | - |
| H. discoidea Decaisne
| 1.1 | 0.7 | - |
| H. gigas Taylor
| - | 0.5 | t |
| H. taenicola Taylor
| 0.3 | - | 0.6 |
| H. gracilis Harvey ex J. Agardh
| 2.9 | - | - |
| H. incrassata (Ellis) Lamouroux
| 7.2 | - | - |
| H. cylindracea Decaisne
| 0.1 | nq | nq |
| H. macroloba Decaisne
| t | nq | - |
U. argenteum Zanardini
U. flabellum (Ellis and Solander) Howe
| 0.2 | nq | nq |
| P. nodulosum Blainville
| t | nq | - |
|
| H. hederacea + copiosa (%)
| 73.5 | 80.5 | 12.1 |
|
| Number samples (N)
| 491 | 155 | 14 |
|
|
t = < 0.05%
nq =present but not quantified
a Part of Coral Sea plateau reef complex
b Originally combined - separated on basis of 5% H. copiosa
|
Table lb. Species composition of Halimeda meadows in individual localities throughout the GBR.
Details as in Table 1a.
Far North Section - ffn = Far Far North; pan = Pandora Cay; gdr = Great Detached Reef; bwc = Blackwood Channel; qun = Quoin Island Entrance; hib = Hibernia Entrance; stm = Second 3-Mile Opening; cre = Creech Reef; wwe = Waterwitch Entrance
Northern Section - liz = Lizard Island; ctn = Cooktown Ribbon Reefs
Central Section - bwl = Bowl Reef
Southern Section - swn = Swains Reefs)
|
|
| Species |
Locality * |
|
|
|
ffn | pan | gdr | bwc | qun | hib | stm | cre | wwe | liz | ctn | bwl | swn |
|
| H. opuntia |
- | - | 18.2 | t | 0.3 | 0.9 | 0.9 | - | - | 1.1 | t | - | 0.1 |
| H. hederacea |
81.4 | 64.6 | 56.0 | 51.5 | 28.2 | 43.9 | 36.8 | 100 | 17.9 | 50.0 | 27.8 | 42.2 | 64.4 |
| H. copiosa |
- | 33.3 | 18.1 | 26.8 | 44.1 | 38.7 | 41.2 | - | - | 21.5 | 46.6 | 34.4 | 12.7 |
| H. minima |
- | - | - | - | - | - | - | - | - | - | - | - | - |
| H. micronesica |
- | 1.1 | 1.4 | 3.7 | - | - | - | - | - | - | - | - | - |
| H. distorta |
- | - | - | 14.5 | 15.2 | 1.0 | 11.3 | - | - | 12.2 | 2.3 | - | - |
| H. melanesica |
- | - | - | - | - | - | - | - | - | - | - | - | - |
| H. lacunalis |
- | - | - | 0.1 | 0.4 | 5.3 | 0.1 | - | - | - | - | - | t |
| H. macrophysa |
- | - | - | - | - | - | - | - | - | - | - | - | - |
| H. fragilis |
- | - | 0.2 | - | 0.4 | 1.7 | 1.3 | - | 0.2 | 2.0 | 3.6 | 20.0 | 7.6 |
| H. tuna |
- | - | - | 0.1 | 0.6 | t | 0.2 | - | 8.9 | 3.1 | 13.7 | - | 0.4 |
| H. discoidea |
- | 0.9 | 1.4 | 0.3 | 0.5 | 1.1 | 1.3 | - | 3.0 | 2.6 | 1.4 | 0.1 | 0.4 |
| H. gigas |
- | - | - | - | - | - | - | - | - | - | - | - | - |
| H. taenicola |
- | - | 1.1 | - | - | 2.0 | 0.1 | - | - | 0.1 | - | - | 3.1 |
| H. gracilis |
15.8 | - | 0.5 | 0.7 | 5.7 | 3.9 | 3.6 | - | - | 2.3 | 1.7 | 2.6 | 0.6 |
| H. incrassata |
2.5 | - | 3.0 | 1.6 | 1.4 | 1.1 | 2.9 | - | 69.8 | 3.5 | 2.4 | 0.6 | 9.6 |
| H. cylindracea |
0.3 | - | - | 0.4 | - | - | 0.2 | - | - | - | - | - | 0.2 |
| H. macroloba |
- | - | - | - | - | - | 0.1 | - | - | - | - | - | t |
|
| Udotea |
- | - | 0.1 | 0.3 | 0.1 | 0.2 | 0.1 | - | 0.2 | 1.3 | 0.4 | 0.1 | 0.6 |
| Penicillus |
- | - | t | t | 0.1 | 0.1 | 0.1 | - | - | 0.3 | t | - | - |
|
H. hederacea
+ copiosa (%) |
81.4 | 97.9 | 74.1 | 78.3 | 72.3 | 82.6 | 77.9 | 100 | 17.9 | 71.5 | 74.4 | 76.6 | 77.1 |
|
| N = |
7 | 6 | 26 | 44 | 84 | 33 | 52 | 1 | 8 | 80 | 76 | 36 | 118 |
|
* Localities as shown in Fig. 1
|
The species composition data is summarized on a taxonomic basis in Fig. 2. Reefal and meadow habitats each contained similar numbers of species from the 4 major sections of the genus Halimeda, although section Rhipsalis, which contains the rhizophytic species normally considered typical of soft substrata, was even less well represented in the meadows than on reefs. However, small amounts of H. incrassata did occur in most of the meadows, while in the Second 3-mile Opening locality divers observed an almost monospecific stand of H. cylindracea that covered an extensive area but was not sampled quantitatively. H.cylindracea was otherwise found only very occasionally.
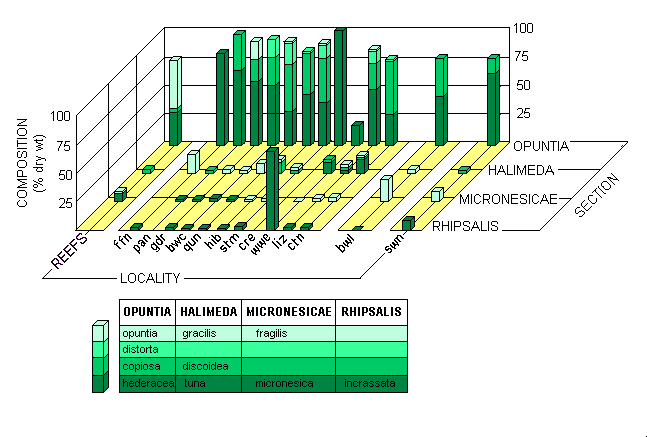 |
|
Fig. 2. Species composition of Halimeda biomass at the 13 localities studied. Locality codes as in Table 1 b
|
The dominant Halimeda species both on the reefs and in the majority of meadows belong to the small-segmented, heavily calcified section Opuntia. Although H. opuntia, a co-dominant species on the reefs, was virtually absent from the meadows, the other major reefal species retained its importance therein. As previously pointed out, Drew (1983) called that taxon H. copiosa, but we now propose to call it H. hederacea. Another important species in the northern and far north meadows, H. distorta, also belongs in section Opuntia. This species had not previously been found on the GBR, but rather as a major component of the luxuriant Halimeda flora on Flinders Reefs, one of the oceanic plateau reefs in the adjacent Coral Sea. There it was co-dominant with H. minima, which did not occur either on reefs or in meadows on the GBR.
H. discoidea, with its large segments and upright habit, was the most visually striking species in the meadows, although, partly because of its very light calcification, it contributed only 1.1 % of the biomass. In addition to 3 other species of section Halimeda, H. gracilis was also found in the meadows, a species not recorded by Drew (1983) but which we have since found on reefs in the Swains Reefs locality in the southern GBR.
The section Micronesicae was represented by H. fragilis and H. micronesica. H.fragilis was found only occasionally on reefs by Drew (1983) but was one of the few species forming the sparse Halimeda flora observed on reefs in the northern GBR (Drew, unpublished). H. micronesica, a species we consider to be a rapid-growing opportunist on reefs, was found, usually as an epiphyte on other Halimeda species, in the Swains Reefs meadows and those in all localities from Quoin Island Entrance northward.
Udotea was as visually striking as H. discoidea in some localities, and small amounts of 2 species, U. flabellum and U. argenteum, were often present together with small amounts of another calcareous green alga, Penicillus nodulosus. These genera were also observed, but not quantified, on the reefs of the central GBR. Because of their structure, they contribute only mud to the sediment, not discrete gravel-sized segments as do most Halimeda species.
RESULTS
Halimeda species--
Species composition--
Species diversity vs depth--
Biomass--
Other algae/seagrasses--
Spatial distribution
Species diversity versus depth
Some Halimeda meadows in the central and southern sections were situated at depths exceeding 50 m, down to a maximum of 96 m. In some situations seabed topography suggested the deep meadows were growing on distinctive mounds of Halimeda debris, as were their shallower counterparts. A marked change in species composition occurred whenever water depth exceeded about 50 m, but there was no significant change in biomass. Relevant data are set out in Table 3. The deeper meadows were characterised by large and heavily calcified plants of H. fragilis and considerable amounts of H. copiosa.
Table 3. Species composition of shallow (<50 m) and deep Halimeda meadows in the central and southern (Capricorn) sections of the GBR.
(Values expressed as % total calcareous green algal biomass)
|
|
| Species |
Locality |
|
|
|
Central
shallow |
deep |
Southern
shallow |
deep |
|
| H. opuntia | - | - | - | 0.5 |
| H. hederacea | 62.5 | 0.5 | 873.8 | 14.7 |
| H. copiosa | 28.9 | 45.6 | 8.8 | 33.6 |
| H. minima | - | - | - | - |
| H. micronesica | - | - | - | - |
| H. distorta | - | - | - | - |
| H. melanesica | - | - | - | - |
| H. lacunalis | - | - | - | - |
| H. macrophysa | - | - | - | - |
| H. fragilis | 3.5 | 53.9 | t | 48.0 |
| H. tuna | - | - | 0.5 | - |
| H. discoides | 0.2 | - | 0.5 | 0.3 |
| H. gigas | - | - | - | - |
| H. taenicola | - | - | 3.7 | - |
| H. gracilis | 3.8 | - | 0.4 | 1.9 |
| H. incrassata | 0.9 | - | 11.1 | 0.8 |
| H. cylindracea | - | - | 0.5 | - |
| H. macroloba | - | - | - | 0.1 |
|
Udotea | 0.2 | - | 0.7 | 0.1 |
| Penicillus | - | - | - | - |
|
| Biomass (gm2 | 179.9 | 57.6 | 389.8 | 184.5 |
| (272.9) | (62.7) | (420.2) | (317.1)
|
| | | |
|
| N = | 17 | 19 | 79 | 39 |
| P = | ---------- 8% ---------- | --------- >1% ---------- |
| ---------------- >>1% ----------------- | |
|
RESULTS
Halimeda species--
Species composition--
Species diversity vs depth--
Biomass--
Other algae/seagrasses--
Spatial distribution
Biomass within the meadows
The quantities of calcareous green algae found at the localities studied are shown in Table 2a. The highest biomass recorded in an individual grab sample was 3157 gm2 dry weight, the highest mean value for all 5 grab samples at a single station was 2081.5 gm2 , and the highest biomass value overall was 4637 gm2 from a diver-collected sample. The mean diver-collected biomass often exceeded that obtained from grabs in a given locality (Table 2b) because diving stations were occupied where grab sampling indicated the presence of a luxuriant Halimeda meadow. Both methods usually yielded similar values when used in close proximity. The depth distribution of calcareous green algal biomass for all the localities is set out in Fig. 3. In the northern sections inter-reefal Halimeda was not found deeper than 48 m; and, indeed, water deeper than 53 m was encountered only in a few narrow canyons penetrating onto the shelf through passages between the outer barrier reefs. Direct comparison with the deep biomass data from central and southern sections is therefore not possible.
Table 2a. Biomass of calcareous algae from all localities studied.
Values represent gm2 for grab stations where at least one sample yielded 100 gm2 or more Halimeda.
Values in parentheses = 1 standard deviation. Locality code as in Table 1
|
|
| | | | | | | | |
| Locality | Overall mean | Maximum grab | Mean grab | Mean diver |
|
| |
| station | individual | locality | |
|
| ffn | - | | 12.7 | 63.3 | - | | - | |
| pan | 378.8 | (51.1) | 414.9 | 1018.8 | 378.8 | (51.1) | - | |
| gdr | 613.8 | (255.7) | 1039.2 | 1116.0 | 603.2 | (278.5) | 677.4 | (341.1) |
| bwc | 523.7 | (538.4) | 1641.6 | 2828.2 | 523.7 | (538.4) | - | |
| qun | 942.7 | (888.0) | 2108.0 | 3157.0 | 695.3 | (409.4) | 2387.4 | (1570.8) |
| hib | 345.2 | (232.1) | 859.4 | 1080.6 | 345.2 | 232.1 | - | |
| stm | 817.8 | (852.0) | 1609.5 | 2414.6 | 642.8 | (543.6) | 2385.2 | (1384.7) |
| cre | - | | 4.1 | 12.3 | - | | - | |
| wwe | 213.4 | | 213.3 | 391.6 | 213.4 | | - | |
| liz | 593.1 | (497.2) | 2081.5 | 2749.9 | 625.2 | (429.9) | 47.8 | (22.7) |
| ctn | 737.3 | (730.1) | 1281.2 | 1917.4 | 575.3 | (404.4) | 2079.1 | (1187.8) |
| bwl | 134.6 | (116.9) | 360.9 | 974.8 | 134.6 | (116.9) | - | |
| swn | 388.1 | (289.6) | 967.0 | 1606.2 | 354.7 | (274.0) | 806.3 | (181.1) |
|
| Mean | 593.1 | (605.7) | | | 507.7 | (405.5) | 1461.6 | (1313.0) |
| N = | 134 | | | | 122 | | 12 | |
|
Table 2b. Comparison of biomass values for diver and grab stations in close proximity.
Details as in Table 2a. P = probability ex Students' t-test.
|
|
| | | | |
| | | | |
| Locality | Diver biomass | n | m2 | Grab biomass | n | m2 | P |
|
| gdr | 677.4 | (341.1) | 8 | 0.7 | 1939.2 | (103.6) | 3 | 0.3 | 10% |
|
| qun | 3955.1 | (758.3) | 12 | 2.8 | 1686.6 | (1240.1) | 5 | 0.5 | 0.1% |
| 1932.9 | (565.1) | 7 | 2.6 | 568.7 | (199.2) | 5 | 0.5 | 0.1% |
| 650.5 | (190.9) | 9 | 2.2 | 644.4 | (141.0) | 3 | 0.3 | >10% |
|
| stm | 2905.0 | (959.3) | 8 | 2.7 | 1609.5 | (710.9) | 5 | 0.5 | 4% |
| 306.1 | (90.8) | 2 | 0.8 | 393.8 | (149.3) | 3 | 0.3 | >10% |
|
| ctn | 1853.6 | (395.4) | 3 | 1.2 | 617.4 | (369.2) | 5 | 0.5 | 1% |
| 3314.9 | (557.9) | 2 | 2.0 | 1311.9 | (53.3) | 2 | 0.2 | >5% |
|
| swn | 822.0 | (144.5) | 10 | 2.9 | 967.0 | (295.4) | 5 | 0.5 | >10% |
| 790.7 | (219.1) | 10 | 3.1 | 394.6 | (377.7) | 5 | 0.5 | 4% |
|
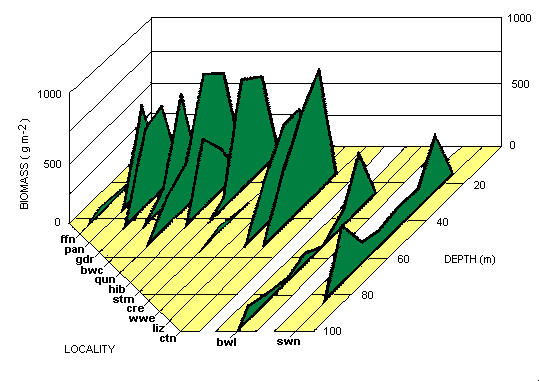
|
|
Fig. 3. Distribution of Halimeda biomass with depth at the 13 localities studied. Locality codes as in Table 1 b
|
Substantial Halimeda-dominated biomass occurred in all localities except the northernmost (far far north), and the overall mean biomass for the meadows, calculated from the 122 grab stations wherein at least one grab sample yielded 100 gm or more biomass, was 508 gm2 total dry weight. As 51 % of all the individual grab samples were devoid of any Halimeda, this biomass value must be regarded as a minimal estimate. Calculations based on the 498 individual grab samples that contained at least a trace of Halimeda resulted in a mean biomass of 579 gm2. These biomass values represent the entire dry weight of the calcareous algae, most of which contained more than 90% ash (Drew and Abel 1985). Using data for the combined species composition of all the Halimeda meadows occurring above 50 m depth, the mean organic content of this Halimeda vegetation was calculated to be 6.0% of total dry weight. This indicates an absolute maximum organic biomass of 278.2 gm2 ash-free dry weight near Quoin Island Entrance, and an overall mean of 30.5 gm2 for all the Halimeda meadows combined.
RESULTS
Halimeda species--
Species composition--
Species diversity vs depth--
Biomass--
Other algae/seagrasses--
Spatial distribution
Species composition - other algae and seagrasses
The data set out in Table 4 show the occurrence of macroscopic marine plants other than Halimeda, Udotea and Penicillus both in and near Halimeda meadows. Stations with at least one grab sample containing at least 100 gm2 dry weight of Halimeda have been considered part of the meadows. Dense growth of large plants of several species of Caulerpa sometimes obscured the understory of Halimeda, particularly in the Cooktown and Swains Reefs localities, while other siphonous green algae such as Struvea elegans and Tydemania expeditionis occurred occasionally within the meadows. With the exception of frequent epiphytic growths of a reddish cyanophyte, probably a Lyngbya spp, on the Halimeda, other algae were not usually found in grab samples from the meadows, nor were they noted by divers. However, a dense growth of the phaeophyte Lobophora variegata was observed to be competing strongly with the Halimeda at one station in the Cooktown locality, while farther north, near Quoin Island Entrance, an apparently unattached Chaetomorpha "bloom" was observed festooning a luxuriant meadow. The seagrasses Halophila decipiens, Hal. ovalis and Hal. spinulosa were occasionally found in meadows, but only the last species occurred in any profusion, and then only in the Cooktown locality.
Table 4. The occurrence of plants other than calcareous green algae in and near the Halimeda meadows.
Upper line = within meadows; lower line = outside meadows
|
|
| | | | | | | | | | | | | | |
| Plant | Locality
|
| ffn | pan | gdr | bwc | qun | hib | stm | cre | wwe | liz | ctn | bwl | swn |
|
Green algae
|
Caulerpa brachypus Harvey
C. cupressoides (Vahl) C Agardh
C. mexicana Sonders ex Kuetzing
C. peltata Lamouroux
C. racemosa (Forsk.) J. Agardh
|
|
-
- |
+
- |
+
+ |
+
- |
+
- |
+
- |
+
- |
-
- |
-
+ |
+
- |
+
+ |
-
- |
+
+ |
|
Struvea elegans Boerg.
|
-
- | -
- | -
- | -
- | -
- | -
- | -
- | -
- | -
- | -
- | +
- | -
- | +
+ |
|
|
|
Tydemania expeditionis A. and E.S. Gepp
|
-
- | -
- | -
- | -
- | -
- | -
- | -
- | -
- | -
- | -
- | +
- | -
- | -
- |
|
|
|
Chaetomorpha spp.
|
-
- | -
- | -
- | +
- | +
- | -
- | -
- | -
- | -
- | +
- | -
- | -
- | -
- |
|
|
|
Monostroma spp.
|
-
- | +
- | -
- | +
- | +
- | -
- | -
- | -
- | -
- | -
- | -
- | -
- | -
- |
Brown algae |
|
Lobophora variegata (Lamouroux) Womersley
|
-
- | -
- | -
- | -
- | -
- | +
- | +
- | -
+ | -
- | -
- | -
- | +
- | -
- |
Red algae |
|
Vidalia spp.
|
-
- | -
- | -
- | -
- | -
- | -
- | -
- | -
+ | -
- | -
- | -
- | -
- | -
- |
Seagrasses |
|
Halophila decipiens Ostenfeld
|
-
- | -
- | -
- | +
+ | -
- | -
- | -
- | -
- | -
+ | +
- | +
+ | +
- | -
- |
|
|
|
Hal. ovalis (R.Br.) Hook. f.
|
-
- | -
- | -
- | -
- | -
- | +
- | -
- | -
- | -
- | +
- | +
+ | -
- | -
+ |
|
|
|
Hal. spinulosa (R.Br.) Aschers.
|
-
- | -
- | -
- | -
+ | -
- | -
- | +
+ | -
+ | -
- | -
- | +
- | -
- | -
- |
|
The grab samples from stations lacking Halimeda gravel usually did not contain much macro-biota, and the sediments usually were muddy shoreward of the meadows or coral sand between the meadows and the outer barrier reefs. The three seagrasses Hal. decipiens, Hal. ovalis and Hal. spinulosa were frequently encountered, particularly on the 2 to 3 km wide strip of coarse sandy sediments between the Ribbon Reefs and the Cooktown meadows and in similar positions in the Blackwood Channel locality further north. Hal. decipiens and Hal. spinulosa were found at depths down to 75 m and Hal. ovalis to 45 m, but biomass levels were very low. Female flowers were observed on Hal. decipiens obtained from 45 m. The muddy areas in the Creech Reef locality adjoining Princess Charlotte Bay supported a significant amount of a red alga, Vidalia spp, as well as occasional plants of Caulerpa, Halophila spinulosa and even Halimeda discoidea. H. cylindracea plants were occasionally recovered from sandy areas peripheral to the more northern meadows.
RESULTS
Halimeda species--
Species composition--
Species diversity vs depth--
Biomass--
Other algae/seagrasses--
Spatial distribution
Spatial distribution of Halimeda meadows and banks
The tops of Halimeda mounds generally lie 20 to 35 m below the sea surface and would not normally be amenable to aerial reconnaissance. However, the particularly good water clarity that must have prevailed during standard monochrome aerial photography of a 120 km stretch of the far north section of the GBR, between about 12.3 and 13.5o S, has provided detailed photographs of shoaling areas behind some of the outer barrier reefs there. These photos showed a mosaic of dark circular patches of the order of 300 m in diameter.
These shoals were initially interpreted by Hopley (1978) as a drowned karst topography, the circular dark areas presumed to be erosion hollows. However, they correspond closely with the tops of those Halimeda mounds detected at depths down to 25 m on echograms of the area. Below that depth they cannot be distinguished from the black of deeper water. The light areas correspond to unvegetated sediment, which occurs between the mounds and elsewhere on the shoals. The correspondence between the typical smoothly undulating Halimeda mound signature on the echosounder scans (illustrated by Phipps et al. 1985; Wolanski et al., in press) and the extent of shoaling areas on the aerial photographs is shown in Fig. 4, together with details of Halimeda biomass and sediment type at the grab stations. These four features were closely correlated throughout the area illustrated. The echosounding tracks and associated grab sampling stations were made without reference to the aerial photographs and therefore represent a totally independent ground-truthing operation.
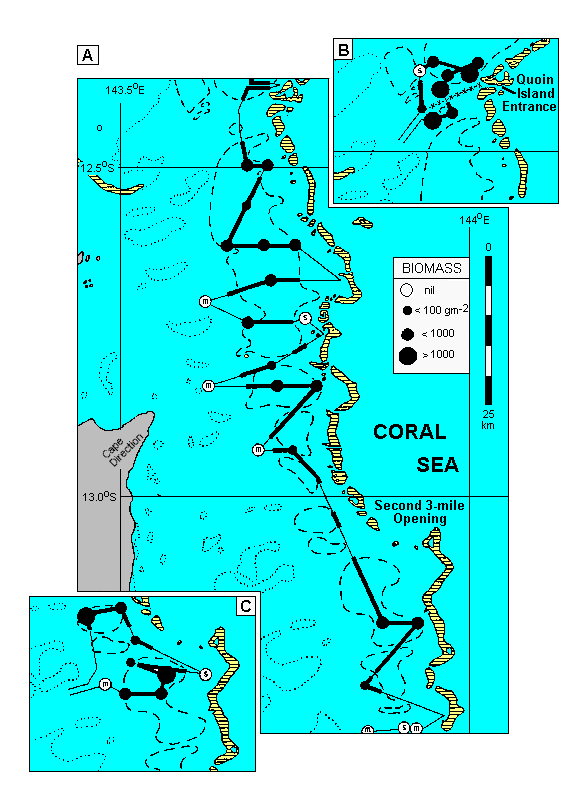
|
Fig. 4. Distribution of shoaling areas, Halimeda vegetation, sediment, and Halimeda mounds
on echograms between 12o 22'S and 13o 22'S.
Insets show data from another cruise to two separate parts of this area.
Dotted lines = reefal areas on mid-shelf (from charts); broken lines = shoals on outer shelf (from aerial photographs); circles = grab stations, diameter indicating biomass; s or m = occurrence of sand or mud rather than Halimeda gravel in grab samples; thick lines = parts of echosounding track with clear signature of Halimeda mounds; thin lines = parts of track without Halimeda mounds; x - x - x - x = track of 80 m deep canyon from Quoin Island Entrance.
The seabed slopes gradually from 0 to 40-50 m depth across the continental shelf, with the reefs and indicated shoals the only significant topographic features. It then drops abruptly to several hundred metres immediately outside the barrier reef tract at the shelf break
|
GO TO Title-
Abstract-
Introduction-
Methods-
Results-
Discussion-
References
Discussion
Observation by divers showed that the Van Veen grab used left a clean, vegetation-free square on the seabed. Although only 0.3 to 0.5 m2 of seabed were sampled at each grab station, the biomass values compared well with samples collected by divers from considerably greater areas, which might have been expected to yield values more representative of the vegetation as a whole. Such a minimum sampling area of about 0.5 m2 is small compared with that of 8.2 m2 determined by Drew (1983) for the very patchily distributed Halimeda vegetation of coral reefs. The homogeneity of the Halimeda meadows was further indicated by the superficial sediments, which were quite consistent within grab stations. Although 1013 grab stations may seem a small number upon which to base conclusions concerning a reef tract extending nearly 2000 km, these stations were concentrated in smaller areas shown by other workers to be appropriate for studies of the Halimeda banks. Their studies, and particularly a survey involving over 6000 stations throughout the GBR reported by Maxwell (1973), made it unecessary for us to consider more than a small proportion of the GBR seabed in the specific context of Halimeda meadows and associated sediment accumulation.
Luxuriant meadows of calcareous green algae, primarily Halimeda species, were found in all sections of the GBR except the far south, where they were not sought in this study because the detailed survey by Maxwell (1968, 1973) found no geological evidence for their existence. The meadows were most extensive from the Cooktown Ribbon Reefs northward to the Blackwood Channel/ Raine Island region. The only area of Halimeda-rich gravel deposits reported in the literature that was not found was that shown by Maxwell (1973) north of Raine Island (our far far north locality), a region that his cruise track only crossed in the south but which we explored more thoroughly. At each of the 13 localities studied some grab stations yielded no living Halimeda or Halimeda-rich gravel sediment, and these were clearly not on Halimeda meadows. However, at a number of stations, small-scale variability resulted in some samples containing no Halimeda, while the others yielded substantial quantities and appropriate sediment. We have therefore considered a station to be on a meadow if at least one grab sample yielded over 100 gm2 living Halimeda and contained appropriate sediment. On this basis 122 (43%) of the grab stations were on Halimeda meadows, and these yielded a mean biomass of 508 gm2 of calcareous green algae, or 593 gm2 if the 76 diver-collected samples are included. The latter were biased toward particularly well vegetated meadows.
The species composition of these inter-reefal meadows had much in common with the lithophytic Halimeda vegetation found on coral reefs, being dominated by similarly sprawling species of the Opuntia section of the genus, and the dominant species were those of the reefs of the GBR itself rather than of the Halimeda-rich oceanic reefs in the Coral Sea. However, there were some significant differences from even the GBR reefs, particularly the almost complete absence of H. opuntia but the presence of H. distorta and H. gracilis. The increased importance of H. fragilis was only in part due to its predominance in the deep meadows, while only a small amount of H. micronesica was found and that mainly in the most northerly meadows, and H. melanesica did not occur at all. These differences cannot be directly attributed to differences in substratum stability since most of the species that were present in significant amounts are themselves normally restricted to solid substrata, according to Hillis-Colinvaux (1980). The Halimeda-rich gravel must offer a sufficiently stable habitat, with the large fragments, often bound together by rhizoids or variously concreted, being quite suitable for colonization.
These luxuriant Halimeda meadows grow on the outer shelf, away from the nutrient cycles of reefs and at depths where the seawater is normally depleted of essential nutrients. However, Andrews and Gentien (1982) demonstrated frequent upwelling of nutrient~rich water from below the thermocline in the Coral Sea onto the deep outer shelf of the central GBR exactly where the deepest meadows occur. That upwelling was generated by internal waves in the thermocline, but would be of insufficient vertical magnitude to lift water over the much shallower sills of the narrow passages between the reefs of the almost continuous barrier farther north. An alternative mechanism, already demonstrated by Thomson and Wolanski (1984) at Raine Island Entrance in the Far North, is the Bernoulli effect, generated by strong tidal currents and perhaps able to lift water over 50 m deep sills. A recent study by Wolanski et at. (in press) in the Cooktown locality demonstrated such upwelling and the subsequent development of a tidal jet, which carried a bottom-tagging layer of upwelled, nutrient-enriched water from a reef passage to the Halimeda meadows. These observations go far toward establishing a causal link between the presence of Halimeda banks and local oceanographic phenomena, and the hypothesis is further supported by the absence of meadows where channel frequency, width or sill depth would be unfavorable for generation of such upwellings. The absence of Halimeda meadows north of the Blackwood Channel locality may therefore be explained by the shallowness of the reef passages there. Veron (1978) indicated that they did not exceed 35 m depth, and Wolanski et al. (in press) showed that nutrients would not be upwelled into a channel shallower than 45 m. The only other significant break in meadow continuity in the northern GBR occurred in the region of Princess Charlotte Bay, where topography still appeared conducive to tidally driven upwelling; but the considerable input of fluvial mud may reduce light penetration too much or directly smother the vegetation. Very muddy Halimeda-rich gravel sediments were associated with the Halimeda meadows at the Second 3-Mile Opening and Waterwitch Entrance localities, which were the nearest to Princess Charlotte Bay although situated 70 km to the north and 90 km to the east, respectively.
Combination of aerial photographs with echograms of the Halimeda banks confirmed that they are composed primarily of mounds about 300 m across, vegetated over much of their surface but with intervening areas of bare sediment (Drew and Abel 1985; Phipps et al. 1985). The photographs did, however, reveal that extensive areas of the banks carried little vegetation, apparently even on the tops of some mounds. Vegetated mounds tended to be concentrated around the perimeter of the banks, perhaps because they have intercepted the intruded nutrient-enriched water first, then either physically prevented its further progress onto the banks or depleted the nutrients before it reached there. Under these conditions, major expansion of the banks may now be lateral rather than vertical.
The sediments associated with these meadows were composed almost entirely of Halimeda fragments, with very little non-carbonate material. The pristine nature of the fragments indicated that they have been generated rapidly, in situ, by death of plants in the overlying vegetation. The sediments had a density of 0.5 - 0.6 gcm3 (Drew and Abel 1985), so accumulation of 1 m of sediment would require 550 kg m2 of calcium carbonate fragments. Wolanski et al. (in press) showed that inter-reefal Halimeda plants photosynthesize at rates very similar to those on reefs, so the productivity rates for reefal Halimeda reported by Drew (1983) can be extrapolated to the meadow vegetation. Those data indicated growth of just over two crops of Halimeda per year, yielding about 1125 gm2 calcium carbonate per year and resulting in accretion of 1 m of sediment in perhaps 500 years. Similar values were calculated by Merten (1971) for shallow-growing monospecific stands of H. macroloba in Guam, where a biomass of about 300 gm2 yielded 738 gm2 calcium carbonate per year, equivalent to 1283 gm2 for the mean biomass of the GBR meadows. At such rates of sediment generation the 10 to 15 m of sediment shown by seismic studies to overlie the Pleistocene discontinuity under these meadows (Orme et al. 1978; Orme 1985; Davies and Marshall 1985) could have accumulated in only 5000 to 7000 years even with little or no sediment input from other sources. Accumulation rates may be twice this amount at localities such as Quoin Island Entrance, with considerably greater biomass. Such rates of vertical accretion are at least as fast as, and probably faster than those of the nearby coral reefs (Davies and Marshall 1985), a possible reason why corals have been unable to gain a hold in these areas.
Data presented by Davies and Marshall (1985), Drew and Abel (1985) and Orme (1985) for both the Cooktown and Lizard Island localities show that Halimeda debris may dominate the sediment of the Halimeda mounds to at least 5 m below their surface, but in most cases this sediment overlies a more muddy sediment, rich in other components such as foraminifera. These deeper sediments lie directly on the Pleistocene discontinuity, itself often mangrove peat, and they must have been laid down earlier in the Holocene marine transgression. This disposition probably occurred in a shallower, more turbid coastal situation, while sealevel was rising rapidly between 10,000 and 8000 years ago. The outer reefs were submerged for much of that time (Davies and Marshall 1985) and the seabed topography was not suitable for generation of strong, localized tidal currents and the resultant nutrient upwellings that are probably necessary for the development of Halimeda meadows, mounds and banks (Wolanski et al., in press). At the start of the transgression, before the reefs were submerged, nutrient upwellings may have occurred and assisted the biogenesis of carbonate-rich sediment, but in situ sediment generation may then have become minimal while the submerged reefs grew upward, reaching the new sealevel only about 6000 years ago. The present-day rates of sediment accumulation calculated above could account for the thickness of the present banks if those rates have been maintained throughout the subsequent 6000 years.
In the Cooktown locality, a submersible survey by Phipps et al. (1985) showed that the tips of Pleistocene coral pinnacles are still visible in the unvegetated hollows between Halimeda mounds. These pinnacles, either still visible or now buried, have not yet been sought on Halimeda banks elsewhere on the GBR, but they may have had a role in controlling the mounded topography of the banks, perhaps by locally modifying water movement or providing shelter for grazing or sediment-disturbing organisms that have prevented Halimeda growth and related sediment accumulation near them. Furthermore, a mounded seabed topography continues several kilometres shoreward of the Cooktown barks. Those mounds do not reach as near the sea surface, are themselves not as thick, and are presently dominated by muddy foraminiferal deposits similar to those underlying the Halimeda mounds. Thus, a reasonably complete successional series representing the mode of formation of Halimeda mounds over many thousands of years may be preserved in the progression from the smaller mounds to those of the Halimeda banks themselves.
Acknowledgements
Shiptime on HMAS Kimbla was generously provided by the Hydrographic Office, Royal Australian Navy, for some of this work. Her master and crew, and also those of the R V The Harry Messel and R V The Lady Basten. provided considerable assistance during field operations. Numerous people took part in SCUBA diving operations, in particular Tony Phipps, Clive Mills, Ray MacAllister and Helen Sturmey, and also in biomass and sediment analysis, particularly Annette Freidl and Wolfgang Falkenberg. Oceanographic ideas were developed in discussions and joint experiments with Eric Wolanski. Peter Davies of the Bureau of Mineral Resources, Canberra. first alerted us to the occurrence of Halimeda banks in the Cooktown locality and also provided vibrocore material from there. David Searle, of the Queensland Geological Survey. alerted us to the existence or the shallow Halimeda banks in the Swains Reefs locality. Taxonomic advice on Halimeda was given by Joanna Noble, Melbourne University, while Ian Price, James Cook University (Townsville), helped with other algae.
<
GO TO Title-
Abstract-
Introduction-
Methods-
Results-
Discussion-
References
References
- Andrews JC, Gentien P (1982)
- Upwelling as a source of nutrients for the Great Barrier Reef ecosystem: a solution to Darwin's question?
Mar Ecol Prog Ser 8:257-269
- Ball SM, Pollard WD, Roberts JW (1977)
- Importance of phylloid algae in development of depositional topography - reality or myth.
In: Frost SH, Weiss MP, Saunders JB (eds) Reefs and related carbonates - ecology and sedimentology. Am Assoc Petrol Geol, Tulsa 4:239-259
- Davies PJ, Marshall JF (1985)
- Halimeda bioherms - low energy reefs, northern Great Barrier Reef.
Proc 5th Int Coral Reef Symp 5: 1-7
- Drew EA (1983)
- Halimeda biomass, growth rates, and sediment generation on reefs in the central Great Barrier Reef Province.
Coral Reefs 2: 101 -110
- Drew EA, Abel KM (1985)
- Biology, sedimentology and geography of the vast inter-reefal Halimeda meadows within the Great Barrier Reef province.
Proc: 5th Int Coral Reef Symp 5: 15-20
- Drew EA, Abel KM (1988)
- Studies on Halimeda. II.
- Reproduction, particularly seasonality of gametangia formation, in a number of species from the Great Barrier Reef Province.
Coral Reefs 6: 207-218
- Goreau T, Graham E (1967)
- A new species of Halimeda from Jamaica.
Bull Mar Sci 17:432-441
- Hillis-Colinvaux L (1968)
- New species of Halimeda: a taxonomic reappraisal.
J Phycol 4:30-35
- Hillis-Colinvaux L (1969)
- Halimeda copiosa and Halimeda hederacea.
J Phycol 5:88
- Hillis-Colinvaux L (1980)
- Ecology and taxonomy of Halimeda, primary producer of coral reefs.
Adv Mar Biol 17: 1-327
- Hopley D (1978)
- Geomorphology of the reefs and reef islands, Great Barrier Reef north of Lizard Island.
In: Workshop on the Northern section of the Great Barrier Reef. Great Barrier Reef Marine Park Authority, Townsville, pp 219-252
- Maxwell WGH (1968)
- Atlas of the Great Barrier Reef.
Elsevier, Amsterdam
- Maxwell WGH (1973)
- Sediments of the Great Barrier Reef province.
In: Jones OA, Endean RE (eds) Biology and geology of coral reefs, vol 1: Geology. Academic Press, New York London pp 299-345
- Merten MJ (1971)
- Ecological observations on Halimeda macroloba Decaisne (Chlorophyta) on Guam.
Micronesica 7: 27- 44
- Neumann AC, Land LS (1975)
- Lime mud deposition and calcareous algae in the Bight of Abaco, Bahamas: a budget.
J Sediment Petrol 45: 763-786
- Orme GR (1985)
- The sedimentological importance of Halimeda in the development of back reef lithofacies, northern Great Barrier Reef (Australia).
Proc 5th Int Coral Reef Symp 5: 31-37
- Orme GR, Flood PG, Sargent GEG (1978)
- Sedimentary trends in the lee of outer (ribbon) reefs, northern region of the Great Barrier Reef province.
Philos Trans R Soc London Ser A 291: 85-99
- Phipps CVG, Davies P.J, Hopley D (1985)
- The morphology of Halimeda banks behind the Great Barrier Reef east of Cooktown, Qld.
Proc 5th Int Coral Reef Symp 5: 27-30
- Thomson RE, Wolanski E (1984)
- Tidal period upwelling within Raine Island Entrance, Great Barrier Reef.
J Mar Res 42: 787-808
- Veron JEN (1978)
- Deltaic and dissected reefs of the far Northern Region.
Philos Trans R Soc London Ser A 284: 23-37
- Wiman SK, McKendree WG (1975)
- Distribution of Halimeda plants and sediments on and around a patch reef near Old Rhodes Key, Florida.
J Sediment Petrol 45: 415-421
- Wolanski E, Drew EA, Abel KM, O'Brien J (in press)
- Tidal jets, nutrient upwelling, and their influence on the productivity of the alga Halimeda in the Ribbon Reefs, Great Barrier Reef.
Est Coastal Shelf Sci
- Wray J (1977)
- Calcareous algae,
Elsevier, Amsterdam